





Why do males and females of many species look so different? Obviously, members of each sex are issued different reproductive equipment to bring together eggs and sperm (collectively referred to as gametes). But what about differences that have nothing to do with an exchange of gametes? Why do male elephant seals have giant blubbery noses while female elephant seals do not? Why does a mallard have a conspicuous green noggin while the female coloration is much better camouflage? Why do males of a species of dung beetle from Ecuador have two horns while the females has none? These types of differences between the sexes are known as sexual dimorphisms (di = two, morph = shape) and Darwin proposed that they arise because of competition for mates.
Natural selection can be broken down into two types of selection. The first leads to differential abilities to survive and is known as mortality selection. The second leads to differential abilities to reproduce and is known as sexual selection. In the preceding chapters, we have talked about adaptations that help organisms survive long enough to reproduce. But surviving to reproductive age doesn’t guarantee you will pass on your genes. Only ten percent of the male elephant seals on a beach will sire pups while the remaining 90% will fail to mate at all. In contrast, the majority of females reproduce. To understand why so many males fail, it is useful to think of mates as a resource. Organisms compete for food, water and space to survive. Why wouldn’t they also complete for the mates required for reproduction? Sexual selection is the process by which variations in sexually dimorphic traits or behaviors give some individuals an advantage in obtaining mates.
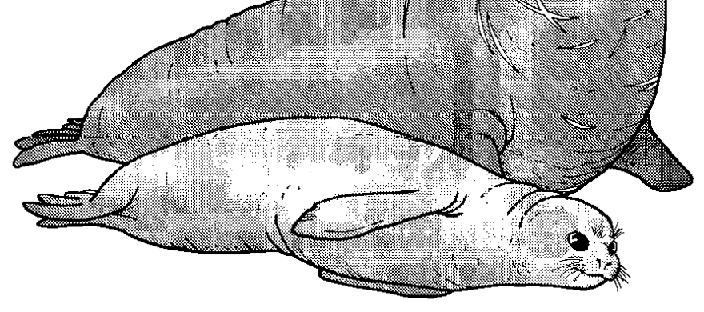
In the case of the elephant seal, females prefer protected areas of beach to have their pups. Consequently, males capable of securing desirable beach front territories will have more luck with the ladies. Not surprisingly, the males that have the most success holding good territories are pretty big. They use their noses to trumpet warnings and threats to other males. This posturing can often dissuade smaller rivals. However, if another male thinks he stands a chance of driving the resident male from his property, the male elephant seals fight. This usually involves these massive animals rearing up and slamming their considerable bulk down upon their opponents while simultaneously driving their daggerlike teeth into their rival’s neck. It can get bloody, but it’s worth it. Successfully holding the territory means holding onto a harem of females with which to breed. So why don’t female elephant seals have big blubbery noses? Or, dagger-like teeth? The answer is, they don’t need them. They aren’t fighting for access to males, the males are fighting for access to the females. But, why fight in the first place? |
 |
 |
|
Figure 4.1 A male (big nose and big mouth) and female elephant seal.
Sexual selection can occur through a number of different mechanisms, but before we address them, we should stop to consider why it happens at all. What drives a system of differential reproductive success? The simple answer is investment. There is usually a disparity between what males and females invest in reproduction.
Neither male nor female loggerhead sea turtles provide any care for their young after the eggs are laid. After they exchange gametes, the male may go off to mate again. The female, however, must expend considerable metabolic energy to build a protective shell for each egg and provision them with the nutrient rich yolk. Then she drags herself out of the water, crawls up the beach, digs a big hole in the sand, lays her eggs (of which there may be hundreds) and covers them up. This is an exhausting process that exposes her to potential harm by terrestrial predators, including humans. Needless to say, if the female is going through all of that, she is going to be choosey about the father of her children!
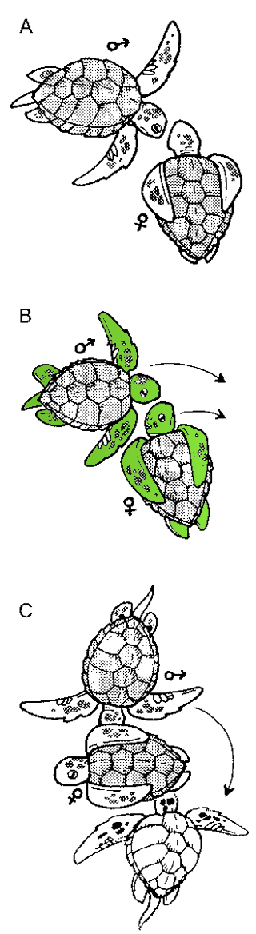
To convince a female that his genes are worth the trouble, the smaller male loggerhead must perform a courtship behavior. He begins by circling the female as she floats facing him (Fig. 4.1 A and B). After circling for awhile, he nuzzles her side (Fig. 4.1C) and bites her back flippers (Fig. 4 .ID). He finishes by facing her and paddling his front flippers in a circular fashion (Fig. 4 .IE). If she likes what she sees at this point, she might accept him (Fig. 4 .IF). Or she might not, in which case the male might keep trying or eventually give up. This type of sexual selection, in which the female selects with whom she will or will not mate, is known as female choice.
What does a choosey female look for in a mate? Sometimes, she’s looking for direct benefits such as she gets from a male with a good territory (as is the case with elephant seals) or a male that will stick around and help with the kids. Those are very tangible benefits. Sometimes, the benefits aren’t as easy to spot. In some cases, females might be reading certain sexually dimorphic traits as indicators of indirect benefits, such as good genes. Female gray tree frogs prefer males that have longer calls. The males themselves don’t offer the female anything directly (such as a good place to live or parental care). However, offspring of males with longer calls grow faster and are bigger when they go through metamorphosis than offspring of males with short calls. So, call length tells the female that the male has better genes for acquiring resources. By choosing males with longer calls, she is equipping her offspring with an adaptation that will give them a distinct advantage as they are competing for food.
Of course, all of this choosing can get out of hand. Runaway selection describes a phenomenon in which female choice rapidly selects for more and more extravagant male features. Male stalkeyed flies with longer eye-stalks tend to be better at acquiring resources. Thus the length of the eye stalk is an indication of good genes and as a result females show a preference for this trait. This preference appears to have a genetic basis and can be passed on to the female’s offspring. Under these conditions, the females of the each generation will preferentially breed with the males with the longest eyestalks. It isn’t hard to imagine how this selection will start to run away with itself and drive eyestalks of males in an population to elongate rapidly. As one might imagine, these flies expend more energy making and carrying around eye stalks and are easier targets for predators than flies without long eye stalks. However, the benefits of winning a mate and passing on one’s genes more than compensate for the inconvenience and expenditure of energy.
There are natural limits to runaway selection. If eye-stalks
became so big that the male could not lift his head, that would
severely curtail his ability to escape predators, find food and mate.
The length of the eye-stalks are probably the result of a balancing
act between sexual selection by females and mortality selection on
males.
I’m a Fighter, Not a Lover!Competition is another way to gain access to mates. This type of sexual selection can take on a number of different forms and can occur before, during or after the fertilization of the egg by the sperm. The most common form of competition is male combat, in which males fight to gain direct access to females or to defend a territory that females find attractive. To be winners, males must have sexually selected traits like bigger bodies or antlers to help them prevail against other males. In stalk-eyed flies, bigger eyestalks correlate with bigger overall size. So, it isn’t surprising that when push literally comes to shove, bigger flies with bigger stalks prevail and get to reproduce. But an exchange of gametes is still no guarantee that one’s genes will get passed into the next generation. Females of many species can mate multiple times before their eggs are fertilized. Sperm competition is the phenomenon in which the sperm from multiple males scramble to get to an egg first. Males can respond to this competition in a number of ways. They can produce more sperm or sperm of varying sizes and shapes that are better adapted to compete in the female’s reproductive tract. Superior number or quality is not the only way sperm competition occurs. In some cases, mating order plays a role. In chimpanzees, the dominant male is often the last to mate with a female and his sperm is more likely to fertilize her eggs. Female insects can actually store a male’s semen in a structure called the spermatheca for extended periods of time before she uses it to fertilize her eggs. In the event she encounters a male that she deems superior to her first mate, she can mate again. At that time, the first male’s sperms can be displaced. Damselfly males have evolved a diverse array of appendages on their penises designed to scoop out a rival’s sperm. To prevent this type of sperm displacement, several species of animals use a techniques called mate guarding. Male loggerhead turtles often stay connected to the female long after the exchange of gametes is complete to prevent another male access. Infanticide is a competition strategy that is sometimes used after offspring have been born. Groups of langur monkeys consist of one male and his harem of females. Male langurs born into the harem must leave. To pass on their genes, these males must eventually depose an older male and take over his harem. When a langur has done this successfully, he will kill his rival’s offspring so that all parental care and resources are invested exclusively in the offspring that he will soon sire.
|
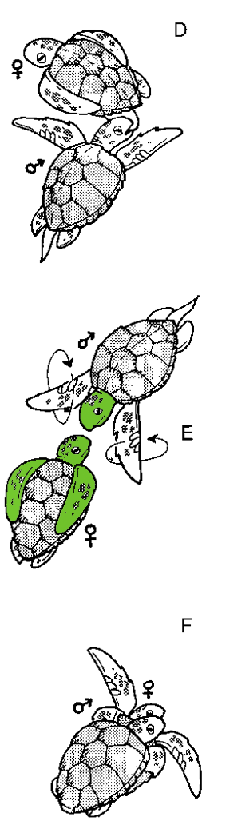 Figure 4.2 Courtship ritual of the loggerhead turtle. See text for description. Modified from Frick, et al (2000). |
|
Sensitive GuysOur discussion, so far, has centered on female choice and male adaptations that develop via sexual selection. But it is best to consider sexual selection in more general terms. Specifically, sexual selection happens when the least-invested sex competes and the most-invested sex is choosey. When sexual selection occurs in nature, it is generally the females that are most invested (for many of the reasons outlined above), but there are occasions when this role is reversed. Males of the firefly Photinus offer females a nuptial (or mating) gift called a spermatophore. The spermatophore is a gelatinous package composed of sperm and food for the developing eggs. During the breeding season, each Photinus firefly will mate several times. However, they do not eat during this time. So, each mating event diminishes their reserves and they aren’t replenishing themselves with food. Consequently, over the course of the season, females become dependent upon the spermatophore’s provisions for their developing eggs. At the same time, the male’s spermatophores are becoming smaller and less abundant. Thus, at end of the season, female-female competition arises for spermatophores and males become selective in their choice of mates. |
 Figure 4.4 Numerous species of dung beetles have evolved horns. Depending on the species, those horns can develop on A) the front of the head, B) the back of the head or C) the thorax. However, when resources are allocated to building horns during development, fewer resources are available for neighboring organs such as eyes. |
||
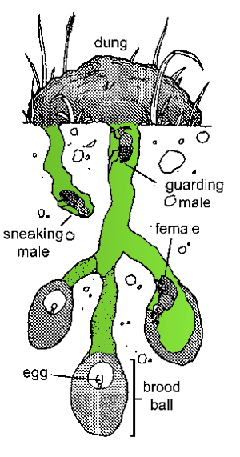
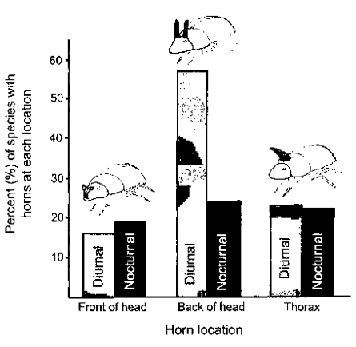
Tag-Team SelectionThere comes a time in every young dung beetle’s life to seek a mate. At this time, males and females head for the best pile of poop they can find. The female burrows under the dung and builds a network of chambers (Fig. 4.3). Each chamber will contain one egg and a provision of dung for the emerging beetle larva. While the female is working below, male dung beetles fight for an opportunity to mate with her. This is done by standing at the mouth of the burrow and defending it against other males. Males that successfully defend the burrow can periodically go down and mate with the female. As it would happen, males with bigger horns are better at securing a burrow than smaller males with small horns. Dung beetles belong to the genus Onthophagus, in the scarab beetle family. This family contains in excess of 2000 species living in a variety of habitats across the globe. (Poop is everywhere, after all.) Most of the Onthophagus beetles have horns, but their location on the body varies from species to species. Members of some species have horns growing from the front of their head; others have them growing from the back of their head; and still others have them growing out of their thorax (Fig. 4.4). These horns are sexually selected traits that are energetically costly to make and haul around. And, since they are part of the beetle’s exoskeleton, they can’t be shed at the end of the breeding season as deer shed their antlers. They also come with a developmental cost. Resources that are allocated to grow horns are not available for building other nearby structures. Thus, beetles with horns growing from the front of the head have smaller antennae than those without horns in the front. Those with horns at the back of the head have smaller eyes, and those with horns on the thorax have smaller wings. This system is a fascinating example of the interplay between sexual selection and ecology. Horns are clearly something useful for combat and access to females, but they come at a cost. Consequently, mortality selection should select for the horn location that has the least impact on the species’ ability to survive. This appears to be the case. Dung beetles active during the day have horns on the back of the head much more frequently than nocturnal species (Fig. 4.5). Bigger eyes are an advantage when trying to navigate the low-light conditions at night. Thus, though sexual selection may favor the evolution of horns in dung beetles, mortality selection would favor nocturnal beetles that grow them somewhere that doesn’t make their eyes smaller.
|